1. Stem cell biology of spermatogenesis 2. Molecular basis of spermatogenesis |
Å@
1. Stem cell biology of spermatogenesis 2. Molecular basis of spermatogenesis |
![]()
| 1. Stem
cell biology of spermatogenesis
Contact address |
| 2. Molecular basis of spermatogenesis | |
i) Monoclonal antibodies
Contact address |
|
| ii) Cloning of testicular germ
cell-specific genes
Contact address |
|
| iii) Haploid cell biology of
spermatogenesis
Contact address |
|
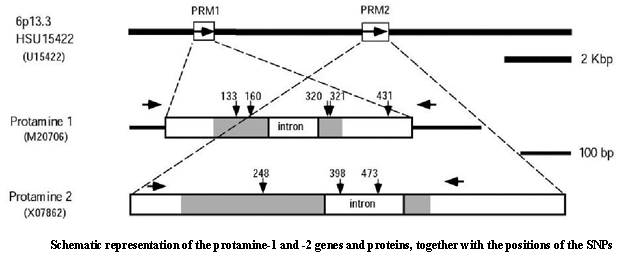
3. Male infertility and SNP analysis in humans More than one tenth of human couples suffer from infertility, and half of these cases are attributable to deficient spermatogenesis in males (McLachlan et al. 1998). Exposure to artificial chemicals is increasingly common, and some of these may act as endocrine disrupters (ED), causing defects in spermatogenesis leading to infertility (U.S. Environmental Protection Agency 1998). Although the molecular basis of most human spermatogenesis disorders remains unclear (Johannes 2000), we hypothesized that deterioration in the effect of male germ-cell-specific genes gives rise to male infertility. Indeed, targeted disruption studies of germ-cell-specific genes have provided us with promising leads to further our understanding of the male infertility mechanism. For example, calmegin, which is a germ-cell-specific molecular chaperone that we cloned, produces infertility in male mice owing to the loss of sperm adhesion to the egg by gene disruption (Ikawa et al. 1997). Our new strategy is to compare the DNA sequences of fertile and infertile men. Since some germ cell-specific genes are intronless, it is easier to examine SNPs or detect mutations by direct DNA sequencing of the PCR-amplified products of chromosomal DNA from blood samples (Figure).
Contact address |
| 4. Applications of the results of our study | |
| i) Diagnosis of male infertility and
Development of novel treatment From SNPs analyses
of many germ cell-specific genes in male infertile cases,
we can identify genes whose function loss results in male
infertility. Developing new ways of detecting SNPs in
these genes should be used as a diagnostic tool for male
infertility. |
|
| ii) Development of new
contraceptive methods A better approach to
develop new contraceptive ways may be to focus on the
development of male contraceptives without any effects on
hormonal functions. For this, it would be better to
disrupt spermatogenesis at the late stage. The
identification of genes specifically expressed in haploid
spermatids and the characterization of these gene
functions would help us to develop new methods of
contraception. |
|
| iii) Development of tests for
the toxicity of drugs, foods, and environmental
disruptors Toxicity tests are an indispensable
step in the development of new drugs. We propose studies
to detect toxicity not only in somatic cells but also in
germ cells using the results of molecular biological
analyses of germ cell-specific gene functions. These
studies will look at the impairments that cause germ cell
infertility or mutations in order to determine possible
serious side effects that may be passed on to progeny. |
|
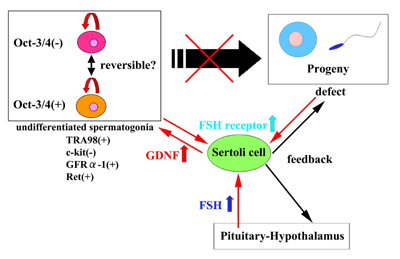 Understanding and
controlling the mechanisms regulating stem cell
proliferation and differentiation are very hot topics in
developmental biology and stem cell medicine. However,
the mechanism of stem cell self-renewal and
differentiation remains elusive, because proliferation
and differentiation occur simultaneously and are
difficult to analyze. Testes with defective germinal stem
cell (GSC) differentiation are a useful model for
studying the behavior and properties of GSCs (Nishimune
and Okabe, 1993). Although no specific molecular marker
for GSCs has yet been identified, we recently
demonstrated that GSCs are part of the undifferentiated
spermatogonial population (c-kit-, TRA98+) by germ cell
transplantation (Ohta et al. 2000). Some mutant testes
deficient in germ cell differentiation have a plentiful
supply of undifferentiated spermatogonia containing GSCs.
Therefore, GSC proliferation can be investigated
independently from differentiation. The undifferentiated
spermatogonial population in deficient testes had Oct-3/4-positive
and -negative spermatogonia, and proliferation was highly
accelerated (Tadokoro et al. 2002). The proliferation of
undifferentiated spermatogonia was regulated by GDNF
induced by follicle-stimulating hormone (FSH). Our data
suggest that the GDNF/FSH pathway controls GSC
proliferation to maintain the total population of GSCs
and their progeny (Figure). Understanding the differences
and similarities between GSCs and somatic stem cells is
one of the most important issues in stem cell biology.
Understanding and
controlling the mechanisms regulating stem cell
proliferation and differentiation are very hot topics in
developmental biology and stem cell medicine. However,
the mechanism of stem cell self-renewal and
differentiation remains elusive, because proliferation
and differentiation occur simultaneously and are
difficult to analyze. Testes with defective germinal stem
cell (GSC) differentiation are a useful model for
studying the behavior and properties of GSCs (Nishimune
and Okabe, 1993). Although no specific molecular marker
for GSCs has yet been identified, we recently
demonstrated that GSCs are part of the undifferentiated
spermatogonial population (c-kit-, TRA98+) by germ cell
transplantation (Ohta et al. 2000). Some mutant testes
deficient in germ cell differentiation have a plentiful
supply of undifferentiated spermatogonia containing GSCs.
Therefore, GSC proliferation can be investigated
independently from differentiation. The undifferentiated
spermatogonial population in deficient testes had Oct-3/4-positive
and -negative spermatogonia, and proliferation was highly
accelerated (Tadokoro et al. 2002). The proliferation of
undifferentiated spermatogonia was regulated by GDNF
induced by follicle-stimulating hormone (FSH). Our data
suggest that the GDNF/FSH pathway controls GSC
proliferation to maintain the total population of GSCs
and their progeny (Figure). Understanding the differences
and similarities between GSCs and somatic stem cells is
one of the most important issues in stem cell biology.
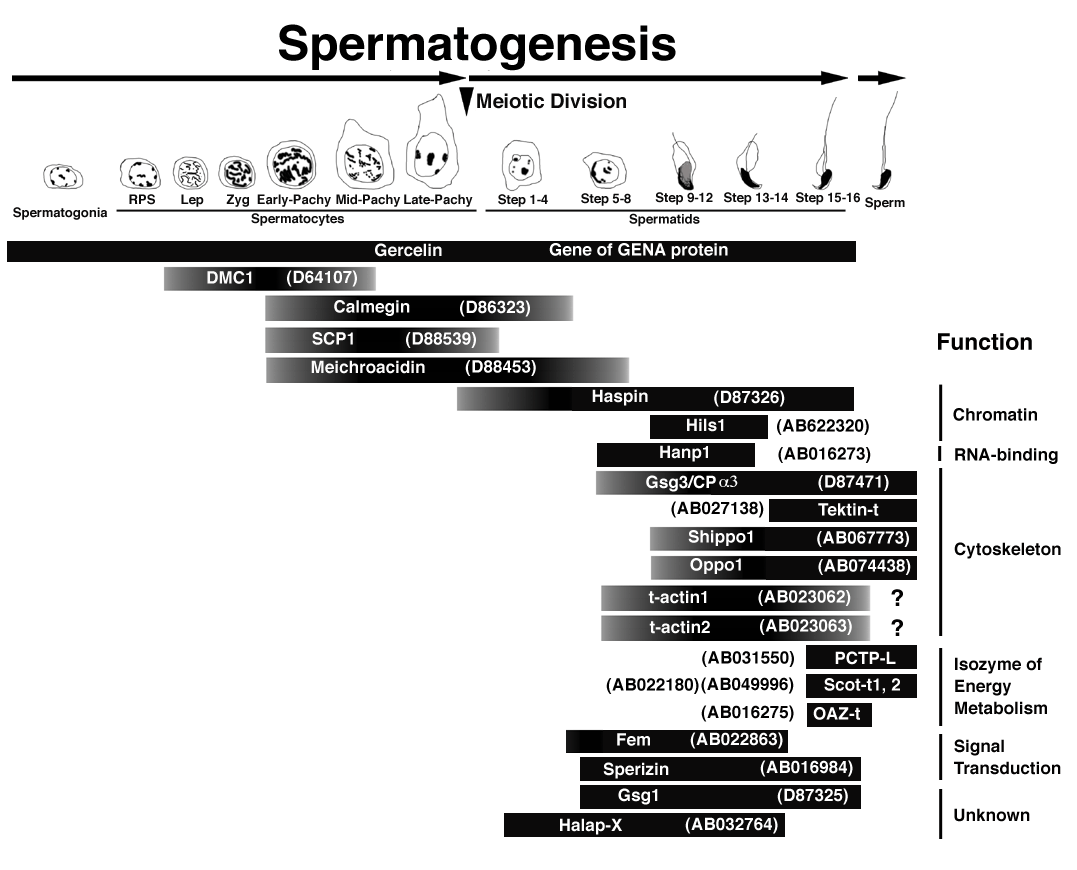
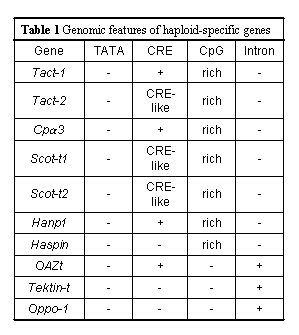 The third process of spermatogenesis,
spermiogenesis, involves nuclear condensation, the
elimination of most of the spermatid cytoplasm, and the
formation of the tail and acrosome. In this phase, more
haploid-cell-specific genes were actively transcribed
than was expected; the expression pattern of each gene
with various putative functions is strictly regulated (Figure
2). Since mammalian haploid cells differentiate into
sperm without dividing after meiotic division, unlike
those in yeasts and some insects (Ross et al. 1993),
there should be some specific regulatory mechanisms
during cell cycle arrest. Elucidating the roles of
haploid-germ-cell-specific genes will facilitate
understanding of not only spermiogenesis but also of the
differences between haploid and diploid cells.
The third process of spermatogenesis,
spermiogenesis, involves nuclear condensation, the
elimination of most of the spermatid cytoplasm, and the
formation of the tail and acrosome. In this phase, more
haploid-cell-specific genes were actively transcribed
than was expected; the expression pattern of each gene
with various putative functions is strictly regulated (Figure
2). Since mammalian haploid cells differentiate into
sperm without dividing after meiotic division, unlike
those in yeasts and some insects (Ross et al. 1993),
there should be some specific regulatory mechanisms
during cell cycle arrest. Elucidating the roles of
haploid-germ-cell-specific genes will facilitate
understanding of not only spermiogenesis but also of the
differences between haploid and diploid cells.