|
研究内容
これまでの研究の流れ
私たちは、免疫応答に必須のCD40シグナルの関連遺伝子を探索する過程で subtraction を行いセマフォリンファミリーに属するCD100/Sema4DのcDNAを単離しました(Immunity 13 : 621-631, 2000)。当時セマフォリンファミリーは神経発生における神経軸索伸長のガイダンス因子とされており、免疫の分野においては未知の分子群でしたが、私たちは、T細胞に高発現する CD100/Sema4D がB細胞及び樹状細胞の活性化を促進する事を明らかにするとともに(Immunity 13 : 621-631, 2000, J. Immunol. 169 : 1175-1181, 2002)、CD100/Sema4D 欠損マウス及び CD100/Sema4D 過剰発現トランスジェニックマウスの作成・解析から、CD100/Sema4D が免疫応答において重要な役割を果たしていることを証明しています(Immunity 13 : 633-642, 2000, J. Immunol. 167 : 4321-4328, 2001)。更にリンパ球が活性化されると膜表面から生理活性を有した可溶性 CD100/Sema4D が遊離すること、T細胞依存性抗原で免疫したマウスや、自己免疫疾患発症 MRL/lpr マウスの血清中に可溶性 CD100/Sema4D が著しく増加していることから、可溶性 CD100/Sema4D が生体内の免疫反応に関与していることを示しています(Blood, 97 : 3498-3504, 2001)。これら一連のCD100/Sema4Dの機能解析からセマフォリンの免疫系での役割が明らかとなりました(Trends Immunol., 22 : 670-676, 2001)。
次に私たちは CD100/Sema4D 以外の免疫系で発現するセマフォリンを探索する過程で2002年に樹状細胞の cDNA ライブラリーから Sema4A をクローニングし、Sema4A がT細胞の活性化に寄与する事を見出しました (Nature, 419 : 629-633, 2002)。更に Sema4A の生体内での役割を解析する目的で Sema4A 欠損マウスを作成し、Sema4A が抗原特異的T細胞 priming のみならず、Th1/Th2 制御においても重要な役割を担っていることも示す所見も得ております (Immunity 22 : 305-16, 2005)。
上記の CD100/Sema4D と Sema4A の二つのセマフォリンの解析から免疫系で機能する一群の分子群の存在が浮かびあがり、私たちはこれらの分子群を「免疫セフォリン分子群 : Immune semaphorin」を呼ぶことを提唱し、現在ではこの概念は国際的にも受け入れられています(Nat. Rev. Immunol 3 : 159-167, 2003, Adv. Immunol. 81 : 173-98, 2003, J. Cell Sci. 116 : 3463-3470, 2003)。
現在私たちは上記の CD100/Sema4D や Sema4A に加えて、心臓の初期発生や骨の恒常性に関与する Sema6D (Genes Dev. 18 : 435-47, 2004, Nat Cell Biol. 6 : 1204-11, 2004)などのセマフォリンの免疫応答における役割も新たに見出し、これら「免疫セマフォリン分子群」を窓に免疫調節機構の解析を精力的に行っています。
セマフォリンとは

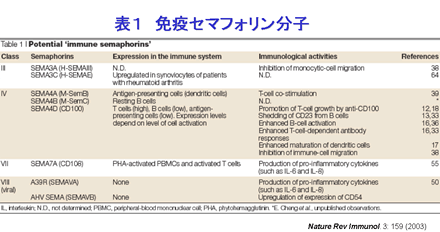
セマフォリン分子群は1990年代に発生過程における神経軸策の方向性を決定する神経ガイダンス因子として同定された分子群ですが、現在その機能は神経系にとどまらず器官形成、血管新生、癌の進展への関与など,多岐にわたることが明らかとなっています。私たちは2000年に免疫系での発現が最初に確認されたセマフォリン分子CD100/Sema4Dの機能解析を通じて,セマフォリン分子が免疫系において重要な役割を果たしていることを明らかにしました。私たちの解析から免疫系におけるセマフォリン分子の重要性が広く認識されるところとなり,現在免疫系において機能するセマフォリン分子は「免疫セマフォリン分子」の名称でも呼ばれています(表1)。
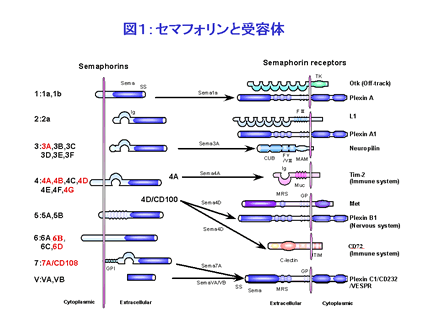
セマフォリン分子は、細胞外にセマドメインと呼ばれるファミリー間で保存された領域を有することを構造上の最大の特徴にしています。現在までの30種類近くのセマフォリン分子が同定されており、セマドメインに続くC末端の構造の違いから7つ、ワクシニアウイルスやある種のヘルペスウイルスなどウイルスにコードされたセマフォリンも含めると8つのサブファミリーに分類されています(図1)。
一方、セマフォリンの生物活性を担う受容体については、神経系ではこれまでに Neuropilin(NP) や plexin ファミリーに属する分子群が報告されています。例えば、Plexin-A、Plexin-B1、Plexin-C1 はそれぞれ Sema1a、Sema4D、Sema7A に結合します。NP-1 と NP-2 はクラスIII型のセマフォリンに結合しますが、そのシグナル伝達は NP と受容体複合体を形成している Plexin-A1 によって担われています。興味深いことに、NP-1 は vascular endothelial growth factor (VEGF) の受容体の一つでもあることが知られています。また免疫系では後で詳述するように、CD72, Tim-2 といった分子、更にはインテグリンに直接結合するセマフォリン(Sema7A)も報告され、セマフォリンの有する多彩な機能を反映してか、非常に複雑なリガンド・受容体関係をとるものと推測されています。
1. Sema3A
Sema3A は神経系におけるガイダンス因子としての機能が世界中で精力的に解析されている代表的なセマフォリン分子ですが、免疫細胞遊走抑制作用が報告されています。また最近種々のヒト腫瘍細胞(胸膜中皮腫、腎がん、肺がん、前立腺がん、大腸がん、乳がん、脳腫瘍など)が Sema3A を分泌していること、Sema3A が MAP キナーゼのシグナル経路を阻止することによりT細胞の活性化を抑制するとの報告がなされ、担がん時の免疫抑制に Sema3A が関与している可能性が示唆され注目を集めています。
2. Sema4A
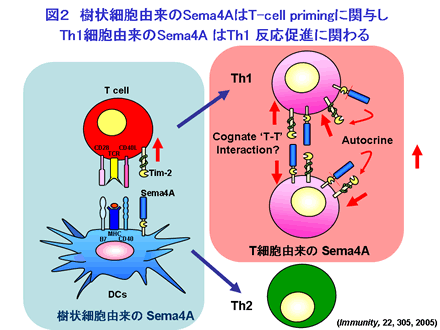
Sema4A は2002年に筆者らが樹状細胞の cDNA ライブラリーからクローニングした分子です。Sema4A は免疫系では主として樹状細胞に高発現しています。休止期のT細胞には Sema4A の発現は認められませんが、Th1 型のヘルパーT細胞に分化するとTh1 細胞特異的にT細胞にも Sema4A の発現は誘導されてきます。私たちはリコンビナント可溶性 Sema4A 蛋白が抗 CD3 抗体で誘導されるT細胞の増殖や IL-2 産生を増強すること、リコンビナント Sema4A 蛋白投与により抗原特異的T細胞のプライミングが増強することから、T細胞活性化における Sema4A の機能を明らかにしています。さらに Sema4A 欠損マウスを樹立し免疫学的解析を行うと、Sema4A 欠損下で、Propionibacterium acnes (尋常性座瘡の原因菌)により誘導される Th1 反応が減弱する一方、Nippostrongylus brasiliensis (糞線虫)感染により誘導されるTh2反応はむしろ亢進するとの知見より、Sema4A の Th1/Th2 制御への関与が明らかになっています(図2)。
3. CD100/Sema4D
CD100/Sema4D はセマフォリンファミリーのなかで免疫系において生理的に発現することが初めて報告されたセマフォリンです。CD100/Sema4D はT細胞に高発現する一方、休止期のB細胞や樹状細胞では発現は弱いのですが、これらの細胞も活性化されると CD100/Sema4D の発現が上昇します。CD100/Sema4D は膜型のセマフォリン分子ですが、リンパ球の活性化に依存して細胞表面で shedding を受け、可溶型としても存在します。可溶型 Sema4D は通常の血清中には検出されないのですが、T細胞依存性抗原を免疫されたマウスや自己免疫疾患を自然発症する MRL/lpr マウスの血清中で検出され、その値は抗原特異的抗体価や自己抗体価の上昇と相関していることから、生理的な免疫反応や自己免疫疾患の進行に関与している可能性も示唆され、生体内での免疫反応をモニタリングするマーカーとしても注目されています。私たちは、CD100/Sema4D 欠損マウスを樹立し免疫学的な解析を行いますと、CD100/Sema4D 欠損マウスではT細胞依存性抗原に対する抗体産生及び抗原特異的T細胞プライミングが顕著に低下していました。また当初は免疫不全を呈する CD100/Sema4D 欠損マウスですが、加齢により自己抗体価の上昇を含む自己免疫疾患が発症することから、CD100/Sema4D が免疫応答及び免疫系のホメオスターシスの維持に重要であることが明らかとなっています。
4. Sema6D
(1) Sema6D と受容体 Plexin-A1
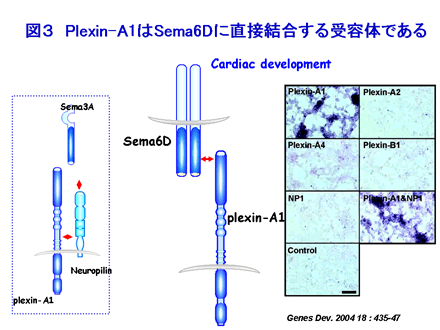
Plexin-A1 は約2000アミノ酸からなる膜蛋白です。1998年分泌型の3型セマフォリン Sema3A が NP-1 と結合すると Sema3A-NP-1-Plexin-A1 受容体複合体を形成し、神経軸索の反兆に関与することが明らかにされ、またニワトリの初期胚の系で6型セマフォリン Sema6D に直接結合する受容体として機能することも明らかとなっています(図3)。
(2) T細胞-樹状細胞相互作用における Sema6D-Plexin-A1 シグナルの役割
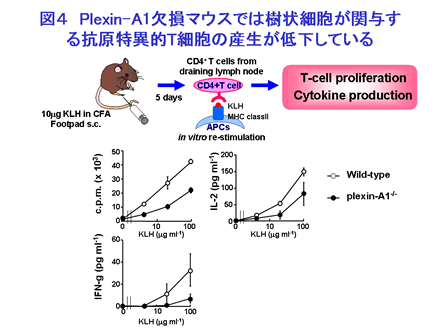
私たちは生体内での plexin-A1 分子の機能を明らかにする目的で、plexin-A1 欠損マウスを作成しその解析を行いました。Plexin-A1 欠損マウスでは、抗原特異的T細胞活性化能の低下を認めるとともに(図4)、Plexin-A1 欠損マウスはヒトの多発性硬化症モデルである EAE モデルに抵抗性であることから、Plexin-A1 が病理的な免疫反応にも関与していることが明らかになっています。また Plexin-A1 欠損樹状細胞では Sema6D の結合低下しており、Sema6D による誘導される IL-12 の産生も低下していることから、Sema6D-Plexin-A1 相互作用の免疫反応における重要性が確認されています。
(3) 骨のホメオスターシス維持における Sema6D-Plexin-A1 相互作用の役割
私たちは、Plexin-A1 欠損マウスの免疫系以外の表現系を調べる過程で、Plexin-A1 欠損マウスにおける大理石骨病の発症を見出しています(図5)。
大理石骨病を発症するメカニズムは骨芽細胞の機能亢進、破骨細胞の機能低下の可能性があるが Plexin-A1 欠損マウスの骨芽細胞に異常は認めなかった。一方 Plexin-A1 欠損マウスでは骨吸収マーカーの値の低下と破骨細胞数の減少を認め、大理石骨病の原因が破骨細胞異常に起因することが明らかとなっている。
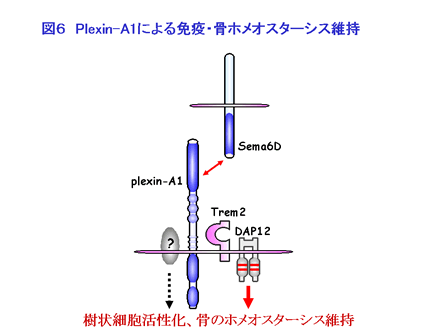
ニワトリの心臓形成期に Plexin-A1 は VEGFR2 や Off-track と心臓の部位特異的に会合して Sema6D の活性を担うことが報告されています。私たちは免疫系においても Plexin-A1 が何らかの受容体分子と受容体複合体を形成して Sema6D の活性を担っているものと考え、スクリーニングを行った結果、Plexin-A1 と Trem-2 (triggering receptor expressed on myeloid cells-2)・DAP12 との会合が明らかになりました。これまでに DAP12 欠損マウスや、ヒトで DAP12, Trem-2 に遺伝子変異を有する Nasu-Hakola 病で大理石骨病の発症が報告されていることから、Plexin-A1 が Trem-2、DAP12 と会合して Sema6D のシグナルを伝え、破骨細胞の機能制御にも関与していることが強く示唆されています(図6)。
5. Sema7A
Sema7Aはヒトの血球上のJohn-Milton-Hagen 抗原として知られていたGPIアンカー型蛋白ですが、データベースを用いた解析からもウイルスセマフォリンの宿主ホモログであることが明らかにされています。Sema7Aのリコンビナントタンパクがヒト単球の炎症性サイトカインの産生、また遊走を促進するという報告からSema7Aの炎症への関与が示されています。またSema7Aは細胞外領域にインテグリン結合モチーフであるRGDモチーフを有するが、神経系での解析でインテグリンに直接結合し嗅神経の伸長に関与することが証明されています。
我々は、Sema7Aが免疫系にも高発現していることに着目し、単球・マクロファージにおけるSema7A受容体の同定を試み,活性化T細胞上に発現するSema7Aが、α1β1インテグリンを介して単球・マクロファージからの炎症性サイトカイン産生を誘導することを見出しました。次にSema7A欠損マウスを用いて生体内での免疫応答におけるSema7Aの役割を検討し、Sema7A欠損マウスでは接触性過敏反応や実験的自己免疫性脳脊髄炎の発症抵抗性を認めています。更に皮膚の接触性過敏反応におけるSema7Aの関与の詳細を検討すると、Sema7Aは局所での炎症反応の誘導に関与していることが明らかになりました。以上のことから,Sema7Aは活性化T細胞上に発現するエフェクター分子であり、α1β1インテグリンを介してマクロファージを活性化することにより、局所での炎症反応促進に寄与していることが明らかになっています。
6. ウイルスセマフォリン
1993年のヒトの Sema3A のクローニングに同時にある種のウイルスにセマフォリン様の蛋白がコードされている事も報告されました。Vaccinia virus により皮膚に炎症が引き起こされる事は良く知られているが A39R はヒトの単球に CD54 の発現を誘導すると共に、IL-6 や IL-8 を始めとする種々の炎症性サイトカインの分泌を誘導することから、Vaccinia virus により引き起こされる病態へのウイルスセマフォリンの関与が示唆されています。また A39R により樹状細胞の機能が抑制されるとの報告がなされ、ウイルス由来のセマフォリンが宿主の免疫系を撹乱する可能性も示されています。
結びに
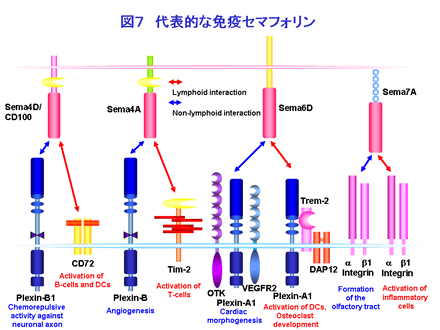
以上、セマフォリン分子の免疫系での役割を最近の報告を中心に述べてきました。現在なされている免疫系におけるセマフォリンの機能解析は主として副刺激分子としての機能に焦点が当てられています(図7)。
しかしながら、セマフォリン分子は従来神経軸索のガイダンス因子として同定されてきた分子群であり,神経細胞の細胞骨格の再編成に深く関与しています。神経軸索の伸長・退縮および細胞の移動という2つの現象は,細胞骨格や細胞接着の変化によってもたらされるという点において共通していることから,セマフォリン分子が免疫系においても免疫細胞移動にも関与している可能性があり、今後免疫セマフォリン分子による免疫制御の研究において,免疫細胞の接着・遊走という切り口での新たな展開が期待されています。
|